| 2011 | 2010 | 2009 | 2008 | 2007 | 2006 | 年 | 次 | 報 | 告 |
| 分子アンサンブル制御・開発研究 Molecular Ensemble Development Research |
|
| 分子アンサンブル測定・解析研究 Molecular Ensemble Analysis Research |
|
|
分子アンサンブル制御・開発研究 局所電子状態,分子間相互作用を設計・制御することによって新しい分子化合物や分子機能を開発することを目指す。大きな目標として,以下の2つのテーマを2本柱とする。 ・分子デバイス実現に向けての基礎の確立 ・触媒機能の制御と高度化(有機金属触媒,タンパク質機能制御)
(1) 二量化が極めて強い金属-dmit錯体における二量体内電荷分離現象の発見 研究担当者:山本(貴),田村,加藤(加藤分子物性研究室) 二量化の強い分子性伝導体は,二量体を1格子点と見なし,1格子点で電荷の二重占有を禁じるクーロン力(Udimer)と,各格子点間における電荷の移動積分(tinter)という,2種のパラメータにて概ねの物性を記述できると信じられてきた。ところが,二量体内電荷分離が観測されないのは,極めて不自然であり,何らかの理由があるはずである。我々は,70 Kで非磁性転移を示す,三斜晶系EtMe3P[Pd(dmit)2]2の分子内振動を測定した結果,転移温度以下で二量体内電荷分離が起こり,二量体2個が繰り返し単位を持つ(四量化)ことが分かった。そもそも,β'型Pd(dmit)2 分子性導体は,二量体が積層構造を持つという構造的特性を有する。従って,格子歪みを伴った四量化と,分子間クーロン力(V)による二量体内電荷分離が共同的に作用できたため,電荷分離が実現したのである。二量体間電荷分離にならなかったのは,(平均)二量体間距離が小さいため,軌道準位の再配列が起こりえないからである(但し,軌道準位の逆転は起こっている)。 観測の結果,四量体の種類は少なくとも2種類あることも分かった。この現象は,イオン性的分子間の二量体間移動積分が大きくなるという四量化と,中性的分子間の二量体間移動積分が大きくなる四量化という,2種類の四量化が共存しており,それぞれに対して,二量体内電荷分離が組み合わされている,ということを意味する。事実,室温から四量化モードが観測されており,これは両者の構造が競合していることを支持している。低温では,四量化モードの揺らぎがほぼ消失したので,どちらか一方の構造が優勢的になっている。この現象は,分子軌道準位の逆転が起こるため,中性的分子間の移動積分の増大が促進されるという,二量体間電荷分離による研究結果と符合する。 一方,二量化の強い分子性伝導体の代表格である,κ-型BEDT-TTF塩は,上記の構造的要請を満たさない。従って,電荷は揺らぐだけに留まり,二量体内電荷分離は起こらないのである。 ところで,β'-型Pd(dmit)2 塩の結晶構造は,積層構造を持たないκ-型BEDT-TTF塩と,二量化が弱く積層構造を持つβ''-型BEDT-TTF塩の中間に位置する。これまでに多く合成されてきた,β'-型Pd(dmit)2の物性は,κ-型BEDT-TTF塩の研究になぞらえた解釈が概ね成り立つ。一方,後者の伝導性は,電荷揺らぎの程度が重要な要因と考えられている。従って,β'-型Pd(dmit)2の中でも二量体内で電荷が揺らぐ物質は,有機伝導体の統一的理解に至るモデル化合物として期待できる。 (dmit = 1,3-dithiol-2-thione- 4,5-dithiolate, BEDT-TTF= bis(ethylenedithio)tetrathiafulvalene) 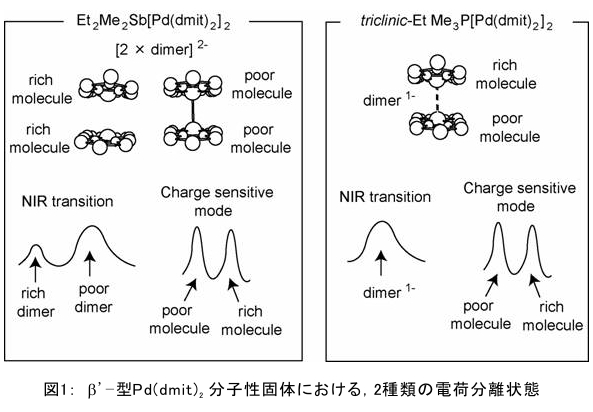 (2) 局在スピンと伝導電子が共存した超分子アニオンラジカル塩 (Me-3,5-DIP)[Ni(dmit)2]2 研究担当者:高坂,加藤,田村,山本(浩),田嶋(陽),深谷(加藤分子物性研究室) (Me-3,5-DIP)[Ni(dmit)2]2は(Me-3,5-DIP)とNi(dmit)2との間に超分子I…およびH…S相互作用を形成したアニオンラジカル塩である。結晶学的に独立な2つのNi(dmit)2アニオン層(Layer I, II)を持ち,Layer Iが局在スピンをLayer IIが伝導電子を持つことがこの塩の最大の特徴である。両者の協奏的な現象を見いだすため,圧力下での電気抵抗測定を行った。静水圧下では,加圧に伴い面内方向で高温領域における金属的挙動が強調されたが,面間方向では絶縁体的な振る舞いが保たれたままであった。低温(約10 K以下)では電気抵抗は各軸方向とも-logT(T: 絶対温度)に比例する振る舞いを示す。面間方向へ一軸性ひずみをかけた時も,電気抵抗の-logT依存性が見られた。これは,近藤一重項のような状態がこの系で実現している可能性を示している。 (Me-3,5-DIP = N-methyl-3,5-diiodopyridinium, dmit = 1,3-dithiol-2-thione- 4,5-dithiolate) 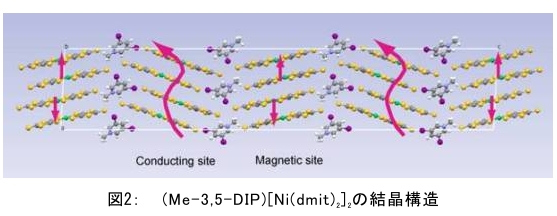 (3) 微小電極による分子性導体の基板上単結晶成長とその電気特性評価 研究担当者:山本(浩),川椙,池田,加藤(加藤分子物性研究室);塚越(河野低温物理研究室) 分子性導体の微小結晶を基板上に成長させ,その電気特性を基板上で直接測定することによって,電界効果・界面現象・結晶サイズ効果等の評価を行った。 (DMe-DCNQI-d7)2Cuのマイクロ/ナノ結晶については, 以前SiO2/Si++基板上で太さ100 nm程度の単結晶について電気抵抗の温度依存性を測定し,バルク結晶で見られる80 Kの金属-絶縁体転移が消失することを見いだしていた。その原因についてはこれまでいくつかの可能性を考え検討を行ってきたが,今回バルク結晶をカーボンペーストとエポキシ樹脂でシリコン基板に固定すると,低温まで結晶が割れることなく測定が可能となり,その場合にも80 Kの金属-絶縁体転移が消失することを確認した。これはシリコン基板の熱膨張係数が数ppm / Kと非常に小さく,その数十倍の熱膨張係数を持つ分子性導体は基板に固定されることによって低温で実効的に負の圧力を感じているためであると考えられる。 (DMe-DCNQI-d7)2Cuに関するこうした解釈に基づくと,α-(BEDT-TTF)2I3で見られた金属-絶縁体転位の転移温度が,基板上のマイクロ結晶では約15 K上昇するというこれまでの結果も,同様の負圧効果が原因であると推測できる。また,転位の温度幅がバルク結晶に比べて広くなる現象も結晶内での歪みが原因であると考えると説明がつく。以上の解釈を前提として,低温の電荷秩序相における電界効果測定を行った。その結果,サンプルはn型の電界効果トランジスタ(FET)動作を示し,I-V曲線はモット・ガーニー則に従うことが明らかとなった。これは一般的な絶縁体にキャリア注入を行う場合と同様の挙動である。昨年度の測定ではON/OFF比が1.1程度と非常に小さい値であったが,今年度は結晶作製条件の最適化を行い,結晶に歪みの生じにくい条件で測定を行うとON/OFF比を40まで向上させることができた。また,基板をシリコンからプラスチックに代えると,転移温度がバルク結晶のそれに近い値となることが明らかになった。一方,α-(BEDT-TTF)2I3は圧力下において,最近注目を集めているグラフェンと同様のゼロギャップ伝導体になることも知られている。ゼロギャップ伝導体は低キャリア密度,高移動度の系であり,電界効果測定に最適の状態にある。従って圧力下での電界効果測定も検討した。現在,シリコン基板を使った実験では圧が均一にかからないが,プラスチック基板を使うと比較的均一に圧力がかかることをつきとめ,さらなる検討を行っているところである。 他方,超伝導体として知られるκ-(BEDT-TTF)2Cu[N(CN)2]Brについても抵抗の温度依存性やゲート電圧依存性について検討を行った。この系は弱い負圧で超伝導体からモット絶縁体になることが知られており,実際シリコン基板上サンプルの抵抗値温度依存性は絶縁体的な挙動を示した。これにゲート電圧をかけるとα-(BEDT-TTF)2I3の電荷秩序相と同様,n型のFET動作をし,ON/OFF比は最大で103に達した。また,サンプルのAFM測定を行い,結晶の厚みを正確に求めるとともに,表面の分子ステップ分布についても観察した。 (DMe-DCNQI = 2,5-Dimethyl-N,N'-Dicyanobenzoquinonediimine, BEDT-TTF = bis(ethylenedithio)tetrathiafulvalene) 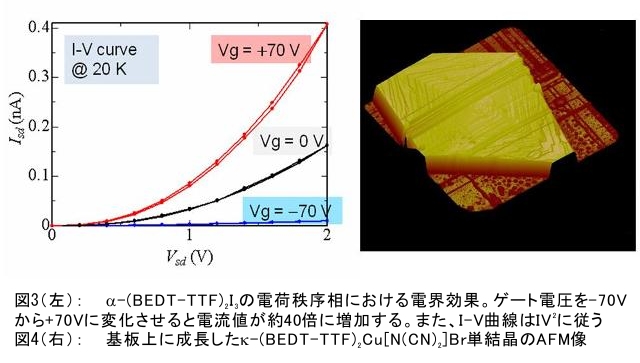 (4) 分子薄膜層における電荷注入と物性制御 研究担当者:川合,加藤(浩)(川合表面化学研究室) 電子デバイスにおける新規な機能を拡充するために,分子性薄膜へ電荷を注入し特性を制御する研究は,未来の分子デバイスに様々な可能性を与えるものと期待されている。本研究では,昨年に引き続き,有機電界効果トランジスタ(有機FET)における電荷注入と電子状態変化に関する研究を進めた。 有機FETは,電界によって有機分子薄膜へキャリアを注入し,薄膜の導電性を制御する電子素子である。基本的な動作原理は,無機半導体と同様に,フェルミ準位近傍の電子状態バンドベンディングによって説明されているが,分子性薄膜の場合,電子状態は局在性が強いため有機FET内の電子状態変化も異なる可能性がある。この違いを解明することは,分子の機能性を生かした未来の分子デバイスを創造する上でとても重要である。本研究では,実際に有機FETを試作して,電荷注入の際に起こる有機分子薄膜内部の電子状態変化を直接観測する手法を確立することに努めてきた。いくつかある手法の中で,最も力を入れているのが蛍光X線を検出するX線吸収分光法(XAS)である。これまでにアセン系薄膜に対して,薄膜内部の電子状態を測れることを確認したほか,全面を金電極で覆った薄膜試料に十分な電界を掛けた状態でも,薄膜内部の電子状態を感度良く測れることを確認することができた。 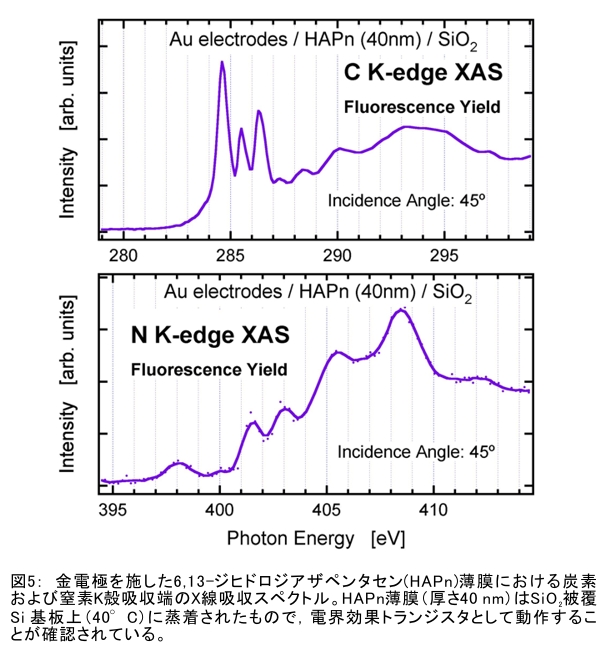 (1) イットリウムポリヒドリド錯体の金属ヒドリド結合へのエチレン挿入に対する計算化学研究 研究担当者:侯,羅(侯有機金属化学研究室) 金属ヒドリド結合へのアルケンの挿入は,水素化反応や重合反応において重要な鍵反応である。このメカニズムに関しては計算化学的手法を用いた単核錯体の研究は数多く行われているが,架橋したヒドリドを有する多核錯体の研究はこれまでほとんど行われていなかった。本年度はイットリウムポリヒドリド錯体[(Me3SiC5Me4)4Y4H8]におけるイットリウムーヒドリド結合へのエチレン挿入反応に対する理論研究を行った。このポリヒドリド錯体には架橋様式が異なるμ4-Hが一つ,μ3-Hが一つ,μ2-Hが6つ存在する。μ4-Hは,4核構造の中心に位置しているためエチレン挿入反応には関与しない。ONIOM(B3LYP:HF)という計算化学の手法を用いてμ3-Hとμ2-Hのエチレン挿入反応について検討した結果,μ2-Hの挿入の方が速度論的に有利になり,μ2で架橋したエチル基を生成することが明らかになった。 (2) 計算化学による希土類ヒドリドクラスターの存在および構造予測 研究担当者:侯,羅,島(侯有機金属化学研究室) 単核のヒドリド化合物は実験および理論計算の面から詳細に研究されている。一方,ヒドリドクラスターは単核の化合物とは異なる物性の発現が期待されるが全く合成例がない。本年度は,Ln3H9やLn4H12等のヒドリドクラスターについて理論計算を行い,これらの存在および構造について予測を試みた。まずLn3H9について3種類の構造異性体を計算した結果,環状の5配位構造が最も安定であることが分かった。予期せぬことに,希土類元素の5d軌道とヒドリドの1s軌道の寄与により,芳香族性を有することが示唆された。またLn4H12については,平面構造や鎖状構造に比べ四面体構造が最も安定であることが明らかとなった。単核の化合物LnH3と比較してLn3H9やLn4H12は,103-204 Kcal/molほど安定であり,これらのヒドリドクラスターが存在しうることを予測した。 一方,イットリウムポリヒドリド錯体とモリブデンヒドリド錯体を反応させることにより,d-f混合金属ヒドリド錯体の合成に成功した。これは,水素と可逆的に反応することが判明し,現在実験および計算の両面からその詳細な反応機構について検討している。 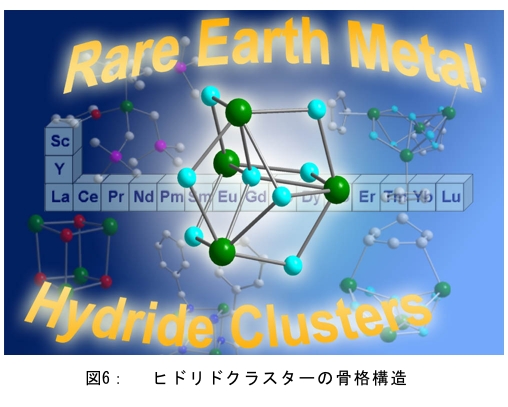 (1) ホスファターゼ阻害剤RK682エナミド誘導体の効率的合成法の確立とVHRとの相互作用解析 研究担当者:平井,小山,袖岡(袖岡有機合成化学研究室) プロテインホスファターゼVHRは,リン酸化セリン,スレオニン,チロシンをすべて脱リン酸化できる両特異性ホスファターゼの一種である。RK682は,長田抗生物質研究室で単離された強酸性の3-アシルテトロン酸骨格を有する天然物であり,顕著なVHR阻害活性を示すことが知られている。我々はRK682を基盤として,中性分子でVHRに対する阻害活性を有するRK682エナミド誘導体(RE誘導体)を開発した。今年度はRE誘導体の効率的合成法を開発し,これまでに比べ大量合成可能なルートを確立した。VHRとRE誘導体との複合体を計算化学的に発生させた結果,RE誘導体はVHRの活性中心に結合していることが示唆されている。現在,この複合体のX線結晶構造解析を長田抗生物質研究室と共同で検討している。 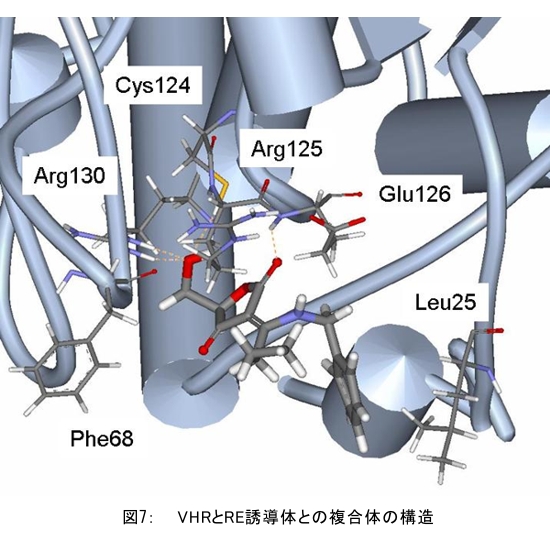 (2) タンパク質と低分子化合物(バイオプローブ)の相互作用解析 研究担当者:奥村,長田(長田抗生物質研究室) グリオキサラーゼI(GLO1)はメチルグリオキサールとグルタチオンからS-D-ラクトイルグルタチオンを生成させる酵素である。これまでに長田抗生物質研究室において,真菌由来化合物ゲルフェリンのメチルエステル体であるメチルゲルフェリン(M-GFN)がGLO1を標的とし,破骨細胞の分化を阻害することを明らかにした。本年度はM-GFNの詳細な阻害機構を明らかにするため,M-GFNとGLO1の複合体のX線結晶構造解析を試みた。マウスGLO1とM-GFNの共結晶化を行い,X線回折データ測定を行った結果,1.7Å分解能で構造解析することができた。得られた構造モデルにおいて,M-GFNはGLO1の基質ポケットに結合しており,M-GFNの有する二つの水酸基がGLO1の活性部位に存在する亜鉛イオンに配位していた。これまでにいくつかのGSH誘導体型阻害剤とGLO1の複合体構造が報告されている。これらの阻害剤は主として亜鉛との配位結合に加え,阻害剤のGSH結合部位がGLO1のアミノ酸残基と水素結合を形成していた。それら阻害剤のうちの一つ,S-(N-hydroxy-N-p-iodophenylcarbamoyl)glutathione (HIPC-GSH)とGLO1の複合体構造においては,阻害剤のGSH部とリジン残基,メチオニン残基,アスパラギン残基,2つのアルギニン残基と水素結合を形成していた。これに対しM-GFNではこの領域において3つのフェニルアラニン残基,2つのメチオニン残基に囲まれ,疎水性相互作用を主とした相互作用をしており,既存のGSH誘導体型GLO1阻害剤とは異なる結合様式をとっていることが明らかとなった。 (1) 2波長同時照射で蛍光シグナルを得られるDronpa変異体 研究担当者:安藤,水野,宮脇(細胞機能探索技術開発チーム) Dronpaは青色光を吸収して緑色の蛍光を発するフォトクロミックな蛍光タンパク質である。490 nmの強い光を照射すると暗状態に移行し,400 nmの照射で元の明状態に戻る。Dronpaにランダムに変異を入れることによって,明状態―暗状態間のスイッチングをより高効率に起こす2つのDronpaの変異体(Dronpa2とDronpa3)を得た。これらの変異体は,アルゴンレーザーの488 nmで励起すると,速やかに暗状態に移行するため,蛍光をほとんど発することができない。一方,紫色レーザー(405 nm)で励起しても蛍光シグナルは出ない。ところが,488 nmと405 nmを同時に照射すると,暗状態―明状態間の往来の結果,明状態の励起分子の数を増やすことができることが分かった。実際に,Dronpa3をHeLa細胞のミトコンドリアと細胞膜にターゲットし,2波長同時照射依存的な蛍光像を撮影することに成功した。 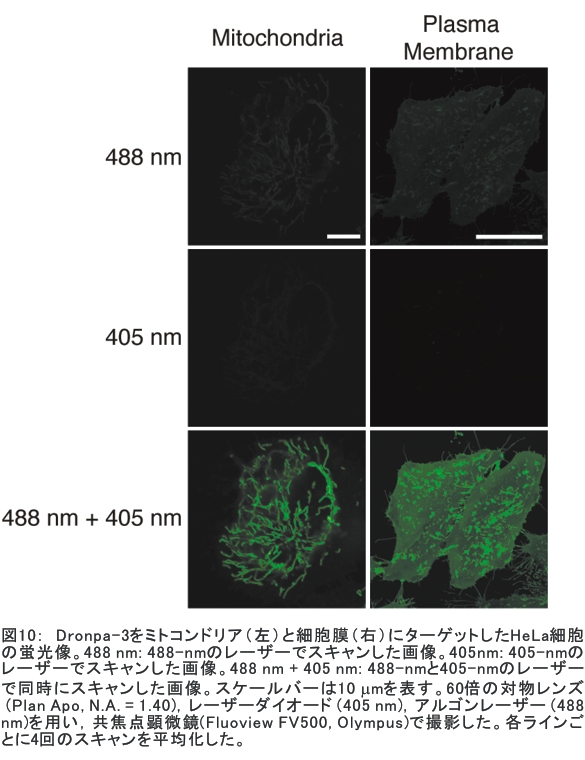 (2) 光異性化能を持つ蛍光タンパク質における構造的なフレキシビリティの光による制御 研究担当者:水野,宮脇(細胞機能探索技術開発チーム) 蛍光タンパク質 Dronpa はフォトクロミズムを示すが,その分子機構は解明されていない。暗状態の結晶構造解析が困難を伴い,明状態の結晶構造だけでは分子機構を議論できないためである。我々はNMRを用いることによって溶液中・室温という条件下での解析を行い,暗状態での構造のフレキシビリティを検討した。光によって発色団とβ-バレルの相互作用の変化が誘導され,これが明状態と暗状態を切り替えていることを見いだした。明状態では,発色団の先端部分が水素結合によってβ-バレルにつなぎ止められ,またβ-バレルから内側へ突き出したイミダゾール環によって発色団の平面性が安定化されていた。これらの相互作用が青い光の照射によって崩され,β-バレルの一部および発色団がフレキシブルになり,無輻射遷移過程が引き起こされていた。 (3) サンゴ由来蛍光タンパク質による凝集体様構造物(dots)の正体 研究担当者:片山,宮脇(細胞機能探索技術開発チーム) DsRed,mRFP1等のサンゴ由来蛍光タンパク質は,哺乳類細胞に発現させた際に凝集体様構造物(dots)を形成することが欠点であると見なされていた。このdotsはアグリソームのように,細胞質における凝集体であると考えられていたが,本研究における検討の結果,リソソーム内腔での蓄積物であること,すなわち,サンゴ由来蛍光タンパク質は酸性条件下でリソソームプロテアーゼに抵抗性であるためリソソーム内腔に蓄積することが示された。また,オートファジー不能細胞でもdotsは形成されることから,細胞質成分をリソソームへ運ぶ未知の輸送機構の存在が示唆されており,この輸送機構については現在検討を進めている。 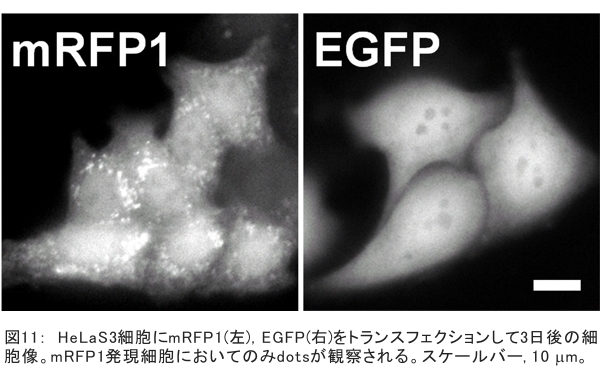 |
|
分子アンサンブル測定・解析研究 測定・解析グループと協力し,広範囲にわたる分子系が示す種々の複雑な現象・機能を局所的電子状態の協奏的連携として理解し統一的原理の構築を目指す。特に,「生体物質の機能の電子論的究明」を大目標の一つとして設定する。
(1) 環境応答型細胞情報伝達系における分子間・分子内相互作用の研究 研究担当者:城,中村,菊地(城生体金属科学研究室) 細菌や菌類,植物の環境(光,酸素,栄養等)感知・細胞内情報伝達は,環境センサーとして働くヒスチジンキナーゼ(HK)と,レスポンスレギュレーター(RR)の二つのタンパク質間のATP 依存性のリン酸基転移反応を介して行われ,「二成分情報伝達系」と呼ばれる。現在,数百種もの二成分情報伝達系遺伝子が明らかになっているものの,HKの環境因子感知の分子機構は依然不明である。環境変化に応答した細胞内情報伝達における,「ドメイン間の分子内情報伝達機構」,「HKのATP依存性自己リン酸化機構」および「HK-RR間におけるリン酸転移機構」を非共有結合相互作用の観点から解明することを目的としている。 (i) 高度好熱菌由来のHK触媒ドメインの高分解能構造を各種条件下で得る事に成功した。十種類の構造比較から,ATP類縁体の結合で,触媒ドメインの基本骨格は変化しないが,その結合部位を覆うループ部分が大きく構造を変化させる事を見いだした。これらの結果と,昨年度構築したHKとRRの複合体構造(3.7Å分解能)を基に,HK自己リン酸化機構との関連で議論した。 (ii) ジフテリア菌はヒト上気道粘膜に感染する病原菌であり,その増殖には血液ヘモグロビンのヘムを主な鉄源として必要とする。ヘムの濃度に依存して,ヘムから鉄を取り出すためのヘム分解系が誘導される事が分かっている。本年度より,このヘム濃度を感知するセンサー系の研究を開始した。ジフテリア菌の二成分情報伝達系ChrS/ChrAがヘム依存的にリン酸転移反応を引き起こす事を初めて示し,この系がヘム分解系の発現を制御するヘムセンサーであると結論した。この自己リン酸化はヘム依存的であり,ポルフィリンや亜鉛,コバルトポルフィリン錯体では効果がないことも判明した。また,ヘムによる活性化にはセンサードメインのHis21が必須の機能を果たしていることが分かった。 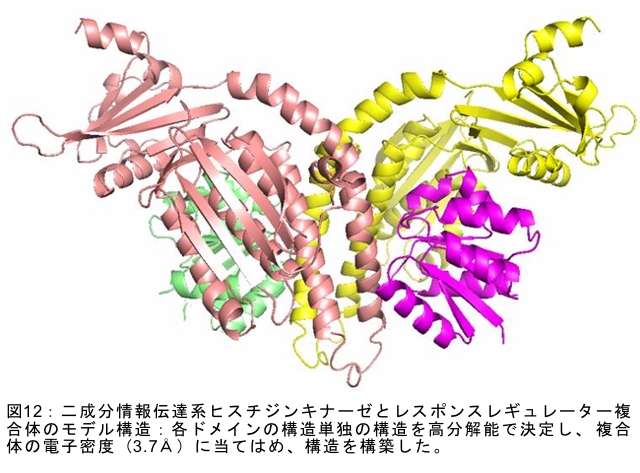 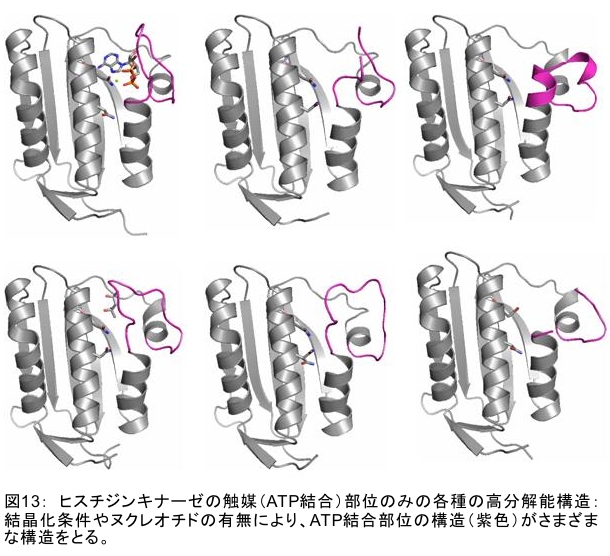 (2) タンパク質-補欠分子構造における分子内相互作用の研究 研究担当者:菊地(城生体金属科学研究室) (i) 人工Croタンパク質の疎水性コアを形成するアミノ酸部位のうち四つを選んで残基置換し,これらの変異体について熱変性実験を行い,協同的フォールディング反応を示す人工タンパク質を得た。NMR構造が決定できるような単一な立体構造の形成は,協同的なフォールディング反応実現のための十分条件とは成り得ず,天然様の協同的なフォールディング特性を実現するためには,さらにアミノ酸配列の絞り込みが必要であることを示した。 (ii) 蛍光を発する状態と無蛍光な状態を可逆的に行き来するGFP様蛍光タンパク質Dronpaの結晶構造を明らかにし,光異性化メカニズムを立体構造ベースで解明することに成功した。 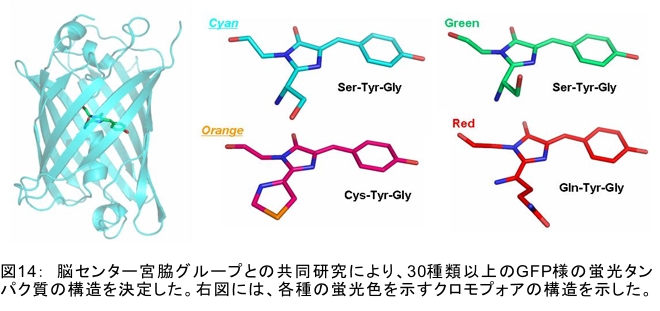 (3) 放射光による分子性結晶ならびにタンパク質の分子内・分子間の電子分布マッピング 高田構造科学研究室では,放射光回折データをマキシマムエントロピー法(MEM)により解析し分子内・分子間の電子分布をマッピングするだけではなく,その電子分布に基づいて得られる静電ポテンシャルも含めて実験的に可視化することにより,構造と分子機能との相関解明を目指している。平成19年度は,研究対象とする物質により研究テーマを以下の3項目に細分化し,個々の研究テーマについて集中的に取り組んだ。それぞれの研究目標を達成するためには,理研内の密接な共同研究が必要不可欠であり,研究テーマiとiiiでは加藤分子物性研究室,テーマiiでは侯有機金属化学研究室と連携して研究を進めた。 (i) 超低温高圧下X線回折による超伝導分子性結晶の構造物性研究とX線磁気回折法の開発 研究担当者:大隅(高田構造科学研究室) 分子性結晶EtMe3P[Pd(dmit)2] 2の超伝導状態の構造研究を目的とした,多重極限条件下X線回折システムの構築を行っている。本年度は,減圧冷却が可能なヘリウム4連続フロー式クライオスタットを導入し,金属ベリリウム製クランプ型セルを1.5 Kまで冷却して試料からの回折スポットが得られることを確認した。今後,クランプ型セルの仕込み圧力の条件探索を行い,超伝導状態にあるEtMe3P[Pd(dmit)2]2の構造研究を目指す。これらの研究に加えて,中性子回折実験が困難な分子性結晶の磁気構造研究を目的として,放射光X線を用いた磁気回折実験の手法開発も行った。  (ii) 異常X線小角散乱法によるタンパク質に収容された金属原子の可視化 研究担当者:伊藤, 水野(高田構造科学研究室) X線小角散乱法は溶液中のタンパク質分子の大きさ,形状,集合状態に関する構造情報を得ることができる強力な手法であるが,異常分散効果を利用した異常X線小角散乱法(ASAXS)により金属含有タンパク質と金属元素の構造情報を分離することができる。さらに,ASAXSによって得られた構造情報を逆モンテカルロ法(RMC)やMEMを駆使して実空間情報へと可視化する技術を合わせて開発し,構造情報と辛グループによる溶液中分子の電子状態に関する情報を組み合わせることにより従来とは質的に異なるタンパク質分子の機能構造相関研究を目指している。フェリチンは分子量48万,内径8 nm,外径12 nmの球殻状タンパク質であり,生体中でFeを蓄積する役割を担っているが,その蓄積過程はまだ解明されていない。また,フェリチンはFeのみならず様々な金属を取り込むことが可能であり,そのサイズをナノレベルで制御できる可能性がありナノテクノロジー分野においても注目されている。本年は,異常分散効果を含んだタンパク質分子のASAXSシミュレータを開発し,金属含有タンパク質中の金属元素の構造情報を抽出できることをシミュレーションにて確認した。また,大型放射光施設SPring-8に設置されている理研専用ビームラインBL45XUにてASAXS実験を行うための測定系の整備を行い,Fe,Ni,CoやAu,Pt等の金属の異常分散効果を測定できることを確認した。現在,表面修飾されたAuコロイド粒子のASAXS測定を行い解析を進めている。 さらに,MEMを単結晶X線結晶構造解析に適用し,金属含有タンパク質の活性中心近傍の電子密度分布を精密に可視化することにより,反応過程を分子構造と電子密度分布の変化として観測する新しい機能相関研究へと発展させることを目指している。本年は,金属含有タンパク質の単結晶X線構造解析データにMEMを適用した際の定量性について詳細に検討した。高分解能回折データが得られているNi-Feクラスターを活性中心に有するNi-Feヒドロゲナーゼでは,MEMにより活性中心周りの電子密度分布の分解能が改善された。構造モデルの精度およびデータの完全性等の問題により活性中心金属の電子数を得るには至らなかった。現在,比較的低分子量の金属含有タンパク質であるミオグロビン等を用いた解析により定量性の向上を進めている。また,侯有機金属化学研究室と連携して,多核希土類ポリヒドリド錯体に対してもMEMを適用し,精密化された電子密度分布から,分子内化学反応のメカニズムを明らかにしようとしている。 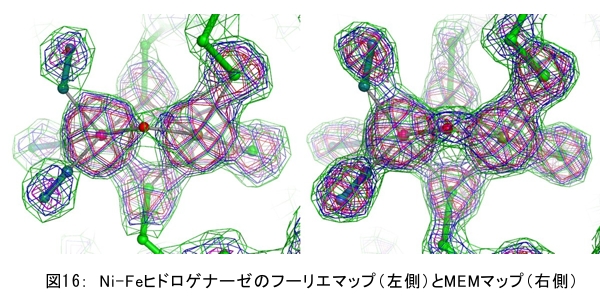 (iii) 電子分布および静電ポテンシャルマッピングによる分子性導体の電荷移動の直接観察 研究担当者:吉田,加藤(高田構造科学研究室) 本研究では,粉末X線回折実験からMEMを用いて電子密度解析を行い,分子性導体α-(BEDT-TTF)2I3の135 Kにおける金属-絶縁体転移の本質についての解明を目指す。平成19年度には,SPring-8のBL02B2にてα-(BEDT-TTF)2I3の粉末X線回折実験を行い,電子密度レベルの構造まで解析可能な信頼性の高い温度変化のデータを30 K,110 K,160 Kの温度について取得することに成功した。得られた回折データに対してMEM/リートベルト解析を行い,相転移前後におけるα-(BEDT-TTF)2I3の電子密度構造を得た。高温相(160 K)と低温相(110,30 K)を比較した結果,ET分子を構成する炭素原子の二重結合に明らかな変化が見られた。二重結合間の結合距離は分子の持つ電荷の大きさを反映していると考えられるため,ET分子間の電荷移動を示唆している。また,セル内の4つの分子について,低温相では電荷の大きさが二つのグループに分けられることが分かり,電荷秩序状態の構造が明らかとなった。現在,電子密度から静電ポテンシャルを計算し低温相における電荷秩序状態の精密解析を進めており,この解析を通じて電荷移動の形態が明らかにされつつある。 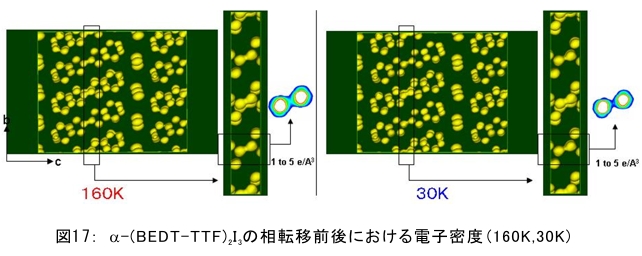 (4) 軟X線発光分光によるヘムタンパク質の電子状態の研究 研究担当者:辛,原田,徳島(励起秩序研究チーム) 昨年度までに,我々の作製した溶液試料用の高分解能軟X線発光分光器を用いて,送液によるヘムタンパク質ミオグロビンの電子状態を観測することに成功したが,軟X線発光分光器の分解能が設計の半分程度に留まっており,スペクトルの微細構造を観測できていなかった。今年度はその原因が表面粗さであることを解明し,分解能があがる回折格子を作製した。今後は,スペクトルの微細構造を議論し,より詳細な理論計算と比較する予定である。これまでミオグロビンの軟X線発光で観測されているd-d遷移はTanabe-Suganoダイヤグラムで説明されることが判明された。また,最低d-d励起は強い直線偏光依存性を示すことを見いだした。この実験結果は,Oh対称性による定性的な解釈ではうまく行かず,歪んだヘムによるD4h対称性で説明が可能になった。その結果,より詳細なクラスター計算によるスペクトルの再現が可能になった。 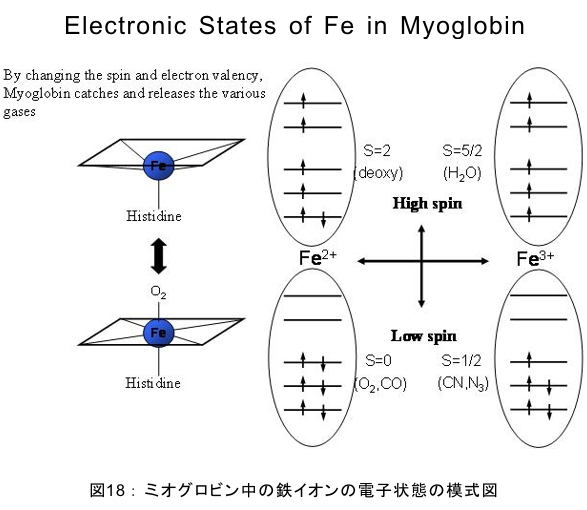 (1) 高分子量タンパク質,タンパク質複合体のNMR解析法の研究 研究担当者:伊藤,美川(城生体金属科学研究室);柴田(柴田上席研究員研究室) 従来のNMRの手法では解析が困難な30 kDaを超える高分子量のタンパク質,およびタンパク質複合体について,構造・機能解析を効率よく行うための新しい測定法・解析法の研究を行っている。平成19年度は,4次元NMRスペクトルを迅速に測定するために,非線形サンプリング法による測定と最大エントロピー法によるデータ処理を組み合わせた方法の研究を行った。4次元NMR法は,曖昧さの少ない解析が可能である等の長所があるにも関わらず,測定時間が長時間におよぶこと,さらに新しい関節観測軸を導入することによる感度低下,間接観測軸のデータポイントの不足によるスペクトルの分解能の問題等から,通常のタンパク質の解析にはこれまで多用されてこなかった。今回開発した方法を用いることで,従来法に比べて非常に短時間で同様の分解能の4Dスペクトルを得ることや,従来法よりもより高分解能の4Dスペクトルを得ることが可能になった。また,細胞内のタンパク質の動態を観測することを目的として,In-Cell NMRの開発研究も継続して進め,高度好熱菌TTHA1718タンパク質について主鎖および側鎖NMRシグナルの帰属を行った。さらにNOE由来の高次構造情報の取得を行うことで,世界で最初の細胞内タンパク質の立体構造解析に成功した。  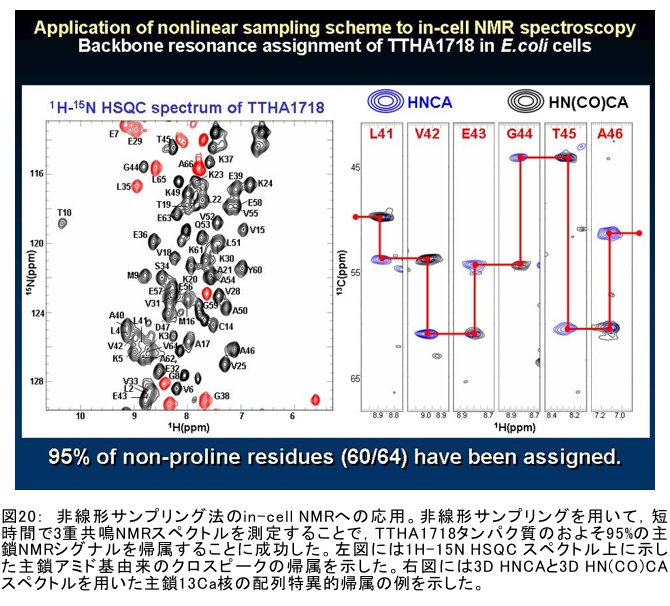 (2) NMR分光法とX線小角散乱を用いたタンパク質間,タンパク質-基質間の相互作用解析 研究担当者:美川,伊藤(城生体金属科学研究室) DNA傷害等の結果生じたssDNA領域はssDNA結合タンパク質(SSB)によって保護され,実際に組換え修復を行うRecAは結合できない。RecF,RecO,RecR等のアクセサリー,タンパク質は様々な相互作用を介してSSBのssDNAからの解離を促し,RecAの組換え修復の進行を助けることが知られている。我々は,NMRと電子顕微鏡観察による相互作用解析の結果,RecOがまずssDNA上のSSBと入れ代わり,次にそのssDNA-RecO-SSBのRecOにRecRが相互作用することによりSSBが部分的に解離することを見いだした。また,RecF,RecO,RecRタンパク質が同時に存在するとRecFR複合体が優位に形成されるが,X線小角散乱による分子形状解析の結果,その複合体はRecF2分子,RecR4分子から なるリング構造であることが明らかになった。リング径やリング内の陽電荷分布からこのリングの内部にDNAが結合することが容易に想像できたため,RecFR複合体はdsDNA上を移動してssDNA-RecO-SSB複合体と相互作用することによりdsDNA-ssDNA領域を認識しているというモデルを提唱した。 (1) STMを利用した単一スピン検出 研究担当者:小野,坪井,花栗,髙木(髙木磁性研究室) スピントロニクスへの期待とともに,局所的なスピン検出への期待が高まっている。局所的な磁気的プローブとしては,走査型ホールプローブ顕微鏡や,走査型磁気力顕微鏡が知られているが,その空間分解能は数μmから数nmにとどまっており,単一原子,単一分子に遡ってスピン検出を行うことは,未だに困難である。単一原子を識別するためにはサブnmの空間分解能が必要であり,そのためには走査型トンネル顕微鏡の利用が現実的である。我々は,磁場中でラーモア歳差運動するスピンとトンネル電流の相互作用を利用する電子スピン回転(ESR)STMの開発に取り組んでいる。本年度はESR-STMの性能テストを行ったが,実験室の音響ノイズが測定に大きな影響与えることが分かり,より静かな環境への装置の移設を行った。また,効率よく測定を行うため,複数の試料と探針を同時に装置内に搬入するための導入室をシステムに付加した。また,ESR-STMとは相補的なスピン検出の手法として,既設の極低温強磁場STMを利用し,電子スピンのゼーマン分裂を非弾性トンネル電流を介して検出する非弾性トンネル分光法を検討している。現在,両手法を用いたスピン検出に適した分子の検討を行っている。  (2) 収量検出磁気共鳴・過渡光吸収検出による2・3 スピン連携 坂口(川合表面化学研究室) 有機半導体では,電子の担い手はラジカルイオンである。ラジカルイオンは奇電子によるスピンを持ち,電極界面では正・負のラジカルイオン対が形成される。このスピン対は一重項または三重項状態で,電荷再結合過程は多重度により変化する。このスピンを磁気共鳴の手法を用いて操作することで,ラジカルイオン対の挙動を制御し,電子移動過程の詳細を明らかにすることができる。 本年度は高分子系有機EL素材である,ポリフェニレンビニレン系の発光挙動に対する磁場および共鳴電磁波の効果を調べた。EL素子を定電流パルスで駆動した場合の磁場依存性は定電圧パルス駆動とほぼ同じで,今後は供給電子数が等しくなる定電流駆動に移行したい。駆動電圧が切れた後の減衰過程に見られる発光強度の磁場効果とマイクロ波効果は通電時より大きく,電場がラジカルイオン間の交換相互作用を増加させていることが分かった。この結果から,有機EL材料の大きな磁場効果の原因は,溶液系で見られる「ラジカルイオン対の寿命が長く,磁場・マイクロ波の効果を受け易い」ためでなく,「電荷再結合が長距離から進むため,交換相互作用が小さい」ためであると結論できる。また,画像計測による研究を開始した。全発光強度で見た場合,高輝度(=高電圧駆動)ほど磁場効果は小さいが,一定電圧で明るい場所と暗い場所には大きな磁場効果の差は見られなかった。 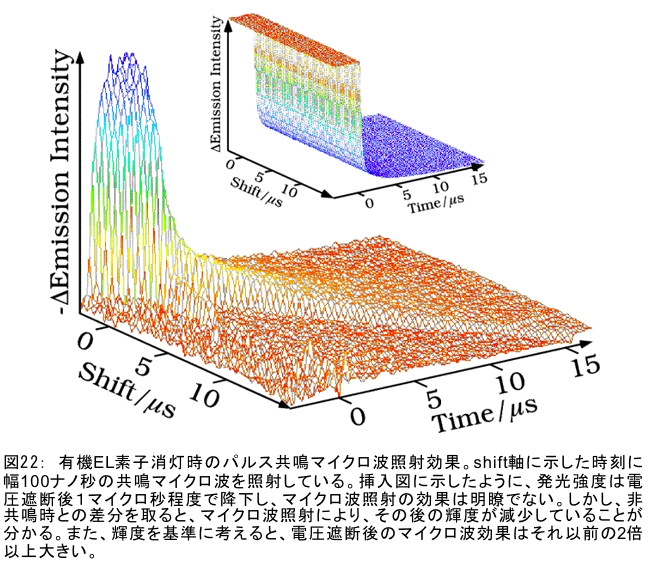 界面選択的偶数次非線形分光の応用 研究担当者:山口,田原(田原分子分光研究室) 昨年度までに,界面選択的なレーザー分光法として二次非線形電子和周波発生(ESFG)分光法と四次非線形ラマン(χ(4) Raman)分光法を開発した。今年度は,(1) ESFGによる気液界面のタンパク質の研究と,(2) フェムト秒時間分解ESFGによる気液界面の分子のダイナミクスの研究を行った。(1)では,気液界面に吸着したタンパク質シトクロムCのESFGスペクトルを測定することによって,タンパク質の表面変性について新しい知見を得た。シトクロムCのSoretバンドのピーク波長は,構造を鋭敏に反映することが知られている。水溶液中(pH=7)で,親水性残基を分子の外側に,疎水性残基を分子の内側に配するように折り畳まれたnativeな構造の場合,Soretバンドのピーク波長は410 nmである。一方,酸性水溶液中(pH=2)で,折り畳まれた構造がほどけた変性状態の場合,ピーク波長は394 nmにシフトする。空気/水界面のシトクロムCのESFGスペクトルは非常にバンド幅が広く,native状態のスペクトルと変性状態のスペクトルを重ね合わせたような形状となった。このことは,気液界面には,折り畳まれたnativeな構造と変性してほどけた構造が混在していることを示唆している。これまでは,気液界面のタンパク質は,疎水性残基を空気側に,親水性残基を水側に配するほどけた構造をとって変性する,と単純に考えられていた。ESFGによって,気液界面のタンパク質を"そのままの状態で"観察することが初めて可能となり,タンパク質の表面変性というよく知られた現象の従来の説明に変更を迫る新しい知見を得ることができた。(2)では,フェムト秒時間分解ESFG分光法を開発して,界面分子のフェムト秒時間分解電子スペクトルを世界で初めて測定した。空気/水界面のローダミン800(R800)の光励起後のダイナミクスは,バルクの水溶液中とは大きく異なることが分かった。励起状態のR800の二量体の解離速度は,気液界面ではバルクの10倍速く,また,バルクでは見られない高速緩和過程が気液界面に存在することが明らかになった。このことは,界面という場の特殊性を端的に示していて,界面特異的な化学反応の探索は非常に有望であることを示唆している。 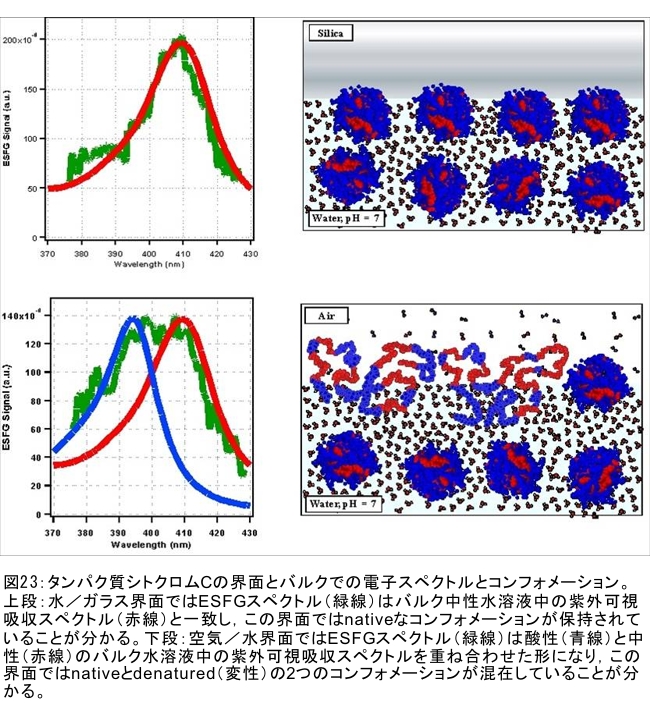 研究担当者:松崎,渡邊(岩崎先端中間子研究室) 理研RALミュオン施設(英国:Rutherford-Appleton 研究所(RAL)内に設置)においては,分子性物質へのμSR測定手法の応用のために,多重極限μSR実験装置を開発中である。この装置は,高圧下測定・微量試料測定・超低温測定・光照射測定等の極限条件下での分子性物質のμSR研究を可能にする。平成19年度においては,この多重極限μSR装置の基幹となる新分光器の開発・製作を進めた。この分光器は,多重極限条件を実現するための十分な空間を確保するとともに,従来のものに比べて3倍以上の600本のミュオン検出器を備える。この分光器を理研RALミュオン施設に設置することにより,微小試料においても高効率・短時間での測定を可能にする。分光器製作にあわせて,検出器系の開発も進めている。これまでにない数の検出器を用いるため検出器自体を小型化するとともに,検出効率を維持するために光波長変換器を利用している。検出器単体の性能試験も実施し,目的性能が達成していることを確認した。 分光器製作と同時に,多重極限条件の一環である高圧下条件の開発を進め,世界でも初の試みとなるHeガスを用いたガス加圧型高圧μSR装置を設計・製作した。このシステムは,試料の設置状況を変えることなく連続的に圧力を変化させることが可能であり,μSR測定を極めて効率的に進めることを可能にするという利点を持つ。RALの高圧グループと共同し,理研RALミュオン施設において得られるパルス状ミュオンビームに特化した高圧セルを製作し,0.64 GPaという設計最高性能の達成を確認した。既に低温での高圧印加試験を終了し,平成20年度からの利用を予定している。 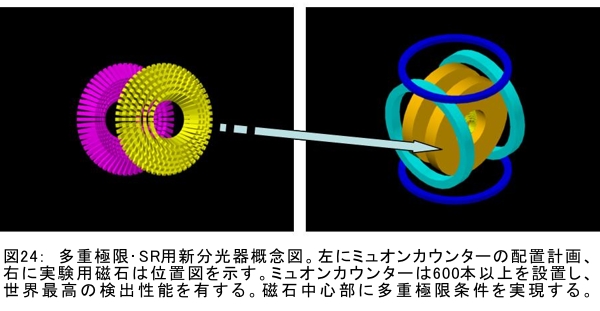 |